
雪真的能把樹木壓矮嗎?
生活在亞熱帶地區的我們,或許對下雪的狀況並不是那麼熟悉。每到冬天,頂多就是有一群人湧向合歡山或其他高山賞雪,體驗一年難得一見的雪景。然而,對於中高緯度地區的人們,下雪可是一種日常現象,冬季的大雪甚至經常帶來各種災害,壓跨建築、影響交通,甚至導致人命財產損失。而且,暴風雪也深刻影響著當地的自然生態,尤其是森林中的植物。樹木被積雪壓彎甚至壓斷的現象並不罕見,讓人好奇的是,如果這樣的雪壓持續作用上千年,甚至上百萬年,是否可能徹底改變樹木的形態?而這種日常現象,是否可能塑造植物的演化?
這個問題乍聽之下似乎有些天馬行空,但在日本的山林中,卻上演了一個真實的案例。故事主角是交讓木(Daphniphyllum macropodum),這是一種廣泛分布於東亞地區的樹木。在日本的暖溫帶輕雪地區,交讓木為高達10公尺以上的喬木。然而,當它們生長於寒冷多雪的冷溫帶地區,就會面對另一套截然不同的生存法則。嚴酷的降雪壓力讓它們不得不「矮化」,轉變為高度不到1公尺的灌木形態。

值得注意的是,灌木型變種的出現,其實並不只是因為雪造成的物理壓力,而是植物在長期適應重雪環境的過程中,透過天擇逐漸演化出的結果。我們可以以交讓木的兩個變種作為例子,即喬木型的交讓木(var. macropodum)以及灌木型的矮交讓木(var. humile)。透過這個例子,我們來探索重雪氣候是如何成為改變植物形態和基因組成的力量,同時深入了解這種適應氣候差異背後的機制。
我們首先來看看這兩個變種的顯著形態差異。喬木型廣泛分布於日本的暖溫帶,這個地區屬於雪量較少的輕雪地帶,這個地區的植株具有直立且高大的樹幹,讓植物可以更有效率效地競爭光照資源。然而,在重雪的冷溫帶,這種形態卻難以生存。在嚴苛的冬季,大量的積雪疊加在高大的樹木上造成樹枝折損,甚至整株樹木都會被雪壓垮。而矮小的灌木型正是適應了這種環境壓力,以低矮的姿態躲過積雪的壓力,更顯靈活地生存下來。
有趣的是,灌木型植物所呈現的橫向生長策略,讓它們能夠以壓條的形式無性繁殖,甚至在持續被雪掩埋的環境中仍然保持良好的族群延續能力。灌木型的形態顯然具有適應性優勢,而這樣「縮身」的樹型應該不僅僅只是一場形態改變事件,更可能牽涉基因層面的遺傳基礎。
那麼,重雪真的能讓植物的基因改變嗎?一項源自於基因組資訊的研究,道出其背後的秘密。
研究表明,喬木型與灌木型之間雖然有遺傳差異,但程度不高,兩者依然有交流。而這種交流的痕跡在兩個變種分布區域相鄰的地區更加明顯。例如,在灌木型分布的南部族群中,研究發現該族群樹高比分布於北部重雪區的族群稍高,推測這很可能是和喬木型的交流後的影響。
儘管有基因交流,灌木型仍與喬木型呈現出令人矚目的遺傳與形態差異。根據分析,兩個變種大約在131萬年前的第四紀中期分開,這個時間正好對應了日本地區重雪氣候開始發展的時期。當時由於冬季風力增強及日本本州山脈的抬升,使得重雪氣候逐漸形成。這樣的古氣候條件下,重雪成為灌木型出現的關鍵天擇壓力,這種能夠適應重雪環境的形態,顯著的提高了存活的機率。
為了要進一步理解這種形態上的差異到底如何與基因關聯,研究者進行了跨地區的種植試驗,將喬木型與灌木型兩個變種同時種植在同樣位於輕雪地區的太平洋沿岸。結果顯示,即使排除了積雪壓力的直接影響,喬木型仍然展現高大直立的樹型,而灌木型依然成長為低矮的灌木形態。這種簡單易懂的試驗清楚地表明,兩個變種的形態差異並不只是因環境直接引起的短期改變,而是由內在遺傳基因所主導的長期分化。
研究者指出,灌木型植株中,與雪壓相關的性狀除了低矮的樹高外,還包括莖的柔韌性提高以及莖的頂端優勢的減弱。這些性狀直接減輕了在厚重積雪下的壓力,讓灌木型得以適應嚴峻的重雪環境。同時,灌木型的橫向生長和無性繁殖策略進一步鞏固了它們在這些環境下的存活能力。
讓我們重新回到最初的問題:「雪是否可以把樹木壓矮?」答案是,可以!但這絕對不是一個單純的物理壓力所造成的現象,而是一個長期的生態適應與演化過程。在重雪氣候的環境中,重雪不僅帶來短暫的結構性衝擊,像是壓斷樹枝或損傷樹幹,還為植物帶來了一種長期的壓力。在這樣的壓力下,只有那些擁有適應特徵的植株才得以存活與繁衍。最終,其中的某些植物演化出了專門適應這種環境的特性,就像矮小的灌木型交讓木一樣。
事實上,這類情形並不只是交讓木的特例。大自然中,相似的現象其實相當常見,例如在面對壓力環境時,許多植物都會展現出形態上的變化。過去,人們經常將這類現象解讀為一種環境影響下的形態塑性(phenotypic plasticity),也就是植物在不同環境下表現出的短期適應能力,而不一定涉及基因層面的差異。然而,這樣的表現是純粹的可塑性反應,還是已經受到基因的深刻影響,隱含著持續的演化過程?這是一個值得深入探討的問題。
例如在臺灣,我們可以觀察到許多長期受盛行風吹襲的地區,其中的森林不僅枝條的分布會明顯偏斜,連整體的高度也顯著下降。這些現象乍看與交讓木的矮化現象相似,似乎也是一種因為環境壓力下的演化結果。然而,這些地區是否也出現類似於交讓木的長期天擇過程,導致植物從形態到遺傳層面都產生了分化?的確是有趣且值得探討的議題。這樣的研究,不僅能讓我們更了解植物如何應對極端條件,也可能幫助我們預測它們在氣候變化下的生存策略。
Yoichi et al. 2023. Genetic differentiation and evolution of broad-leaved evergreen shrub and tree varieties of Daphniphyllum macropodum (Daphniphyllaceae). https://doi.org/10.1038/s41437-023-00637-2

鮭魚之亂—臺灣破傘菊的連續撞名事件
還記得幾年前的鮭魚之亂嗎?當時因為壽司店推出優惠的關係,很多人去戶政事務所把自己的名字改成「鮭魚」,當時的臺灣社會中突然多出了好多鮭魚,也算是蔚為奇觀。人的名字可以因為自己的喜好來取,就算撞名也沒有問題。不過在植物學名中,撞名就是一件重大的事了,「唯一學名」的概念下,後面取的名字和先前的撞名就被迫要換名字,這種現象在植物中其實相當常見,但是像今天的主角這樣連續撞名的,就真的比較少了。今天要介紹的就是臺灣破傘菊的連續撞名事件。
破傘菊屬 (Syneilesis) 是菊科植物中一群很特殊的植物,這些植物通常有掌狀分裂的葉片,葉柄長在葉片中央,形似雨傘一般,中文名稱即因此而來。破傘菊屬也是東亞的特有屬,全屬植物僅有7種,主要分布於中國、俄羅斯、韓國、日本和臺灣。在臺灣,共有兩種破傘菊屬植物,分別是高山破傘菊 (S. subglabrata) 和臺灣破傘菊。這兩種植物都是臺灣的特有種,不過生育地卻天差地別。前者喜歡長在中北部的中高海拔山區,多見於林下透光處,而後者則僅分布於臺灣北部的低海拔地區,長在烈日曝曬的草生地上,兩者的生育地可謂天壤之別。。
過去臺灣的文獻中臺灣破傘菊的學名,常用的是S. intermedia (Hayata) Kitam.,不過也有研究者認為,應該使用的學名是S. hayatae Kitam.,究竟這兩個學名有什麼差別,用哪個才是正確的呢?在釐清這個問題之前,我們先來回顧一下臺灣破傘菊是怎麼被發現的。
臺灣破傘菊最早是由佛里神父 (U. Faurie) 於1903年採集於苗栗 (Bioritsu),隨後在1906年由早田文藏發表為黃菀屬的新種 (Senecio intermedius Hayata),從這之後,臺灣破傘菊的分類地位其實一直都未曾被挑戰過,也就是說,後續的研究都認為它是一個「好種」。不過這個種類的學名,卻莫名的遭到許多的挫折。

故事的起因是早田氏使用的學名S. intermedius,他在發表時,其實未曾注意到已經有個更早的S. intermedius Wight已於1846年被發表了,根據命名原則,早發表的名稱具有所謂的優先權,因此早田較晚發表的名字就撞名,變成後出同名 (later homonym) 的狀況,是不符合法規的名稱。後來,早田或許發現這個種類置於黃菀屬似乎不太對,因此早田氏於1919年又在蟹甲草屬又發表了一個新名稱Cacalia intermedia Hayata,用來取代這個黃菀屬的名稱。
不過呢,這個C. intermedia也遇到同樣的問題,因為早在1891年,Kuntze就已經發表過一個相同的名稱了,所以早田的這個名字又再度因撞名的緣故無法使用。直到1934年,北村四郎 (S. Kitamura) 在〈ヤブレガサ属 (Syneilesis Maxim.) の起用〉一文中,才發現了Senecio intermedius的撞名問題,所以發表了一個替換名 (replacement name) Syneilesis hayatae Kitam.,這才讓臺灣破傘菊終於有了一個合於法規、可用的名字。也就是說,早田氏在兩次的命名中都沒有注意到撞名的現象,但是卻都離奇的撞名了,這種機率到底多高?
不過有趣的是,北村氏又在1937年,根據早田氏發表的C. intermedia ,發表了一個新組合名稱Syneilesis intermedia (Hayata) Kitam.,並且把先前發表的Syneilesis hayatae Kitam. 併到這個名稱中。然而,北村氏可能沒有注意到,其實不管是早田氏發表的哪個名稱,其實都是基於相同的模式標本而來,所以最早發表的Syneilesis hayatae Kitam.才是具有優先權的名稱。
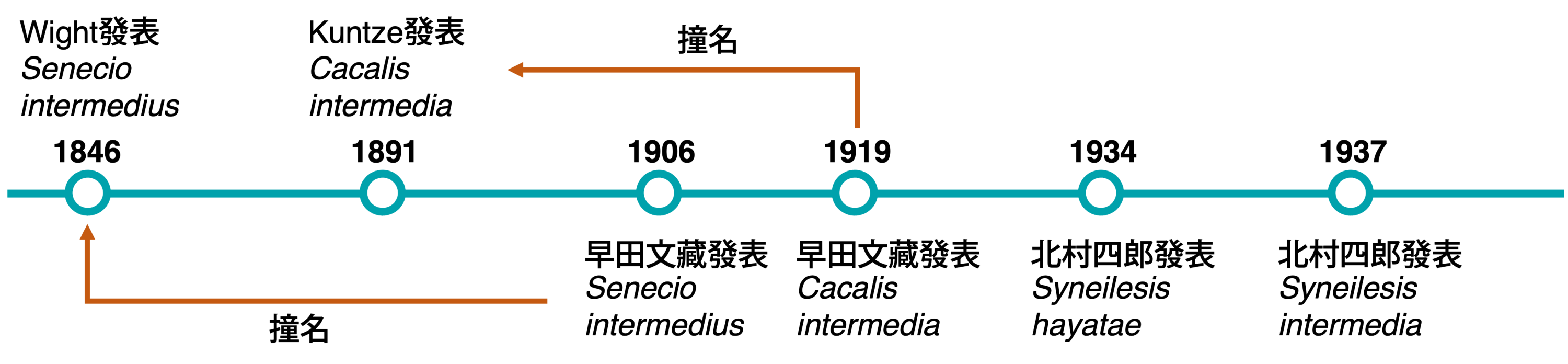
intermedius這個字的意思是在…之間或是中間的意思,可以聯想到命名的人覺得這種植物的形態介於某兩種植物之間 (但常常沒有說明是哪兩種),也因為這種模稜兩可的模糊特性,使得這個字非常常被使用在植物的命名當中,根據Tropicos的紀錄,光是種小名使用intermedius (含不同性別) 的紀錄就有2000筆以上,橫跨18-20世紀,可說是植物分類學史上相當淵源流長的菜市場名了。或許是過於常見的緣故,自1982年以來,就沒有新名稱使用這個字的紀錄了,不禁讓人好奇,進入21世紀後,植物學命名的新「菜市場名」會是什麼字呢?

餐桌上的植物006—甘藍是怎麼變成高麗菜的?
甘藍(Brassica oleracea)是我們日常生活中常見的十字花科 (Brassicaceae) 蔬菜,卻很少有人知道它的身世竟然如此複雜。結球的甘藍(高麗菜)、翠綠的青花菜、皺葉的羽衣甘藍、塊莖狀的球莖甘藍 (大頭菜),這些看似外型各異的蔬菜實則屬於同一種植物。然而,這些蔬菜的祖先是來自於何處呢?這成為科學家長期以來的難題,而近年整合基因體學、考古學與歷史文獻的綜合研究,終於為這道難題提供了新的答案。本文將帶你一同了解甘藍的起源,以及它如何從一種野生植物,透過人類的選擇,成為今日多樣的蔬菜品種。
甘藍的多樣性與人擇的力量
許多人或許都知道,不同品種的狗其實都是同一物種,但你可能沒想到,我們餐桌上的許多甘藍類蔬菜其實也是如此。例如,綠色的青花菜(B. oleracea var. italica)、小巧的抱子甘藍(又稱孢子甘藍,B. oleracea var. gemmifera)、常見的圓形高麗菜(B. oleracea var. capitata)、白色的花椰菜(B. oleracea var. botrytis)、波浪狀葉片的羽衣甘藍(B. oleracea var. acephala)及球莖甘藍(就是大頭菜)(B. oleracea var. gongylodes),它們全都是甘藍。只是在栽培過程中,人們發現,栽培的甘藍很容易產生各種形態變異,例如莖特別肥大的、花朵特別多的、葉片會包起來的,人們刻意保留了想要的特定形態,因此產生了各種不同的蔬菜。早在19世紀,達爾文(Charles Darwin)就曾觀察到甘藍的多樣性,並將其作為人為選擇影響植物形態的一個典範。現代基因體研究也進一步驗證了他的推測。

甘藍的起源來自哪裡?
不過我們吃的這些五花八門的甘藍,到底最一開始是起源在哪裡?有關這個問題,長久以來一直存在不同的假說。有人認為甘藍是起源於西歐沿海地區,因為那裡有一些與栽培甘藍相似的野生種,如被稱為B. sylvestris或B. oleracea var. sylvestris的植物。但也有研究人員提出,東地中海地區,如現今的希臘與土耳其沿海,才是甘藍真正的發源地。
過去由於缺乏足夠的分子資料與可靠的考古證據,因此難以直接證實或否定這些說法。直到近年來,隨著基因體學技術的進步,科學家才能夠透過族群基因體學、親緣分析、生態棲位模擬與考古學方法,對甘藍的起源做出更精確的推測。
來自基因體研究的證據
在一項近期的研究中,科學家分析了224個甘藍品種與其野生近緣種的基因體資料,結合親緣分析與族群遺傳學,試圖找出現存野生甘藍的最近緣祖先。研究結果顯示,愛琴海特有種 B. cretica 是目前最接近栽培甘藍的野生近緣種,而其他過去被認為可能是甘藍祖先的物種,如 B. incana 和 B. montana,則可能是由已馴化的甘藍再次野化而形成的族群,而非原始的祖先。結合考古證據,研究推測甘藍的馴化時間可能最早發生在公元前1250年至1000年之間,地點可能位於現今的敘利亞或東地中海沿岸地區。這些發現否定了將英格蘭或西歐沿海地區視為甘藍馴化中心的假說,進一步支持東地中海才是甘藍真正的馴化發源地。
馴化與野化的循環
研究還發現,甘藍在人類馴化後,仍然能夠「逆向演化」回到類似野生的形態─也就是說,某些栽培品種在特定環境下若不受人工管理,可能會自行恢復成接近野生的特徵,形成新的「野化品系」(feral lineages)。
這種野化現象並不罕見,例如一些在加拿大、丹麥和德國發現的「野生」甘藍,其實可能是栽培甘藍逃逸並重新適應野外環境後產生的族群,而非真正未馴化的祖先形態。這也解釋了為何有些基因分析會顯示這些「野生」族群與栽培品種高度相似。
考古與歷史文獻提供的輔助證據
除了基因組研究之外,考古學與歷史文獻也為甘藍的起源提供了有力的佐證。目前最早的甘藍栽培考古證據來自敘利亞,時間約在公元前1250年至1000年間,儘管更早的遺跡仍有待進一步挖掘與確認。在古希臘約公元前500至200年間的文獻中,已有關於甘藍的記載。例如,植物學之父提奧弗拉斯圖斯(Theophrastus,約公元前320年)曾描述過三種甘藍變種,而古羅馬學者老普林尼(Pliny the Elder)則在其著作中紀錄了至少十種甘藍的類型。這些來自考古與文字紀錄的證據,進一步支持東地中海地區才是甘藍馴化的主要地點,而西歐地區則是在較晚的時期才普遍引入並栽培甘藍。
結語
透過基因體學、考古學與歷史文獻的整合,科學家終於釐清了甘藍的起源—它並非來自英國或西歐,而是源於東地中海的愛琴海地區。B. cretica 被確認為其最近緣的野生近緣種,而馴化過程則發生於公元前1000年前後的東地中海地區。這項發現不僅對演化生物學具有深遠影響,也有助於未來作物改良與保育工作的推動。
Mabri et al. 2021. The Evolutionary History of Wild, Domesticated, and Feral Brassica oleracea (Brassicaceae). https://doi.org/10.1093/molbev/msab183

數位標本影像的應用與極限
植物分類學是一門歷史悠久且講求精確的學科,專門研究植物的鑑定、命名與分類。然而,這門學科的發展長期受限於標本的取得與觀察,特別是模式標本 (type specimen)。傳統上研究人員必須前往標本館或野外調查分析,來取得研究必須的植物材料。近年來,隨著數位化技術的進步,許多標本館開始將館藏標本數位化,讓研究者能夠透過網路取得高解析度的標本影像。這項發展在疫情期間尤顯重要,確保研究工作不會因為封鎖或旅行限制而停滯。
然而,數位標本能否完全取代實體標本?數位影像對於植物分類學究竟帶來多少便利?又有哪些難以克服的限制?在一篇由 Phang、Atkins 和Wilkie 合著的論文中,他們深入探討了這個問題。他們以馬來亞熱帶樹木植物紫荊木屬 (Madhuca)(山欖科)為案例,檢視數位標本在植物分類學上的應用與挑戰。
數位化浪潮下的植物標本館
疫情改變了許多科學研究的運作模式,全球各地的封鎖措施,使研究人員無法親自前往標本館或進行野外調查,因此,線上數位標本的重要性大幅提升。許多植物標本館因此加緊推動數位化,例如荷蘭的 Naturalis、法國的國立自然史博物館(Muséum National d’Histoire Naturelle),以及美國的iDigBio等機構,都建立了龐大的線上資料庫,收錄數百萬張植物標本的數位影像,讓全球研究者得以遠端存取與分析。

目前,JSTOR Global Plants 資料庫等大型數位平台已經收錄超過200 萬份模式標本圖像,大幅提升分類學研究的便利性,並讓全球科學家能夠更容易共享與比對植物標本。此外,一些針對特定植物類群的數據庫,例如 Solanaceae Source(茄科植物資料庫),以及愛丁堡皇家植物園(RBGE)提供的山欖科、秋海棠科和薑科植物資料庫,也成為許多專業分類學研究的重要資源。
「標本數位化」在不同的情境下可能有不同的定義。有些情況下,數位化指的是將實體標本拍攝為高解析度圖像;另一方面,有些標本館則會同時數位化標本的附加資訊,例如標籤上的採集地點、日期、採集者名字及相關分類資訊。部分機構甚至會額外提供色彩校正、比例尺,甚至 3D 掃描技術,以確保標本圖像能夠忠實呈現標本的完整特徵。然而,由於各標本館的資源與技術水準不一,導致數位標本的品質仍存在極大的差異。有些標本的解析度較低,缺乏色彩校正,甚至無法放大觀察細節,這些問題都會影響數位標本在研究中的適用性。
數位影像的應用與侷限
目前,數位影像已能有效支援某些植物分類學的應用,尤其是針對宏觀特徵明顯的植物,例如:
- 物種鑑定:研究者可以透過比對數位標本與相關物種的特徵,判斷植物的種類。
- 模式標本比對:模式標本是某個種類的正式參考標本,數位化後能讓全球研究者遠端檢視,避免運送標本時可能造成的損壞或遺失。
- 確認植物分布:數位標本的相關數據能夠幫助研究者檢視植物的分布範圍及生態環境。
- 形態測量與分析:高解析度的數位影像可以用來測量葉片長度、脈序、果實大小等宏觀特徵,特別是影像包含比例尺時,測量的精度會更高。
然而,數位標本仍然存在許多無法克服的限制。在本研究中,研究團隊發現許多關鍵的微觀形態特徵無法透過數位影像準確辨識,例如:
- 花的細部特徵難以觀察:這些部位往往需要解剖,或使用顯微鏡與微距鏡頭才能清晰觀察。
- 被毛與表皮細節:植物表面的毛、腺體等結構通常需要透過高倍率放大檢視,這是現有數位標本難以拍攝的部分。
- 果實與種子內部構造:如果標本上沒有已經剖開的果實,將無法觀察種子的形態與細部特徵。
研究團隊分析了來自新加坡與柔佛的219 份數位標本圖像,發現只有不到一半的分類學特徵可以透過數位影像準確描述與測量。因此,即使高品質的數位影像能夠協助物種鑑定與分類,仍然無法取代實體標本館的查閱需求。
如何提升數位標本的應用性?
若要讓數位標本更廣泛地應用於植物分類學研究,研究團隊提出以下幾點改進建議:
- 為所有標本圖像加入比例尺:目前許多數位標本缺乏比例尺,導致研究人員無法準確測量形態特徵。未來,應確保所有標本數位化時,皆附加比例尺進行標準化處理。
- 改善光線與對比度:部分標本影像因光線不足,使得葉脈等細微特徵難以呈現,這部分可以透過調整光照條件或後製提高對比度改善。
- 補充微觀形態影像:若能在標準數位影像之外,提供顯微鏡觀察的細部影像,將大幅提升數位標本的分類學價值。
- 整合不同植物標本館的資料:目前不同標本館的數據格式與命名方式不一致,若能透過國際組織(如GBIF)推動標準化,將能讓影像更易於統一比對與應用。
結語
數位標本影像的大規模應用,無疑為植物分類學研究帶來變革,讓研究人員在無法遠行時,也能夠線上取得關鍵性的研究材料。然而,目前的數位化技術仍無法完全取代實體標本,尤其是在需要微觀特徵來區分物種時,實體標本的查閱仍然不可或缺。未來,隨著數位化技術的進一步發展,或許可以透過標準化的數位化流程、高解析度影像、以及 AI 輔助技術,使數位植物標本的應用更加成熟。
Phang et al. 2022. The effectiveness and limitations of digital images for taxonomic research. https://doi.org/10.1002/tax.12767

「解明」鼠尾草的真相
所有喜歡香草植物的人,大概都聽說過鼠尾草,作為唇形科 (Lamiaceae) 中種類最多的屬,鼠尾草屬植物在世界各地被廣泛研究與栽培,涵蓋了約1000個物種,園藝上也培育出相當多的品系。無論是花園裡的觀賞植物、藥用植物,還是自然棲地中的野生種類,鼠尾草其實比想像中的更常見。不過,這個屬的多樣性也帶來了一些有趣的分類學謎題—特別是那些長得「看似一樣」的物種,是否真的就是同一種植物?
本文主角正是一個剛剛從這個龐大的植物家族中被發現的隱藏種 (cryptic species)。經過研究團隊的深入研究,來自羅馬尼亞東南部的這個不知名植物,最終被證明與相似的奧地利鼠尾草 (Salvia austriaca) 截然不同,並被正式命名為解明鼠尾草 (Salvia revelata)。這則分類學的故事,不僅讓我們認識鼠尾草的多樣性,也彰顯了研究者如何在細節中發掘真相。
隱藏物種的起點
故事從羅馬尼亞的一片草原開始。研究團隊在野外調查時,發現了一種不明的鼠尾草,它的整體外型讓人以為就是奧地利鼠尾草,不論是植株高度、葉片的形態,甚至花的顏色,都與其非常相似。如果是平常的觀察者,恐怕沒有人會多看一眼。然而,熟悉鼠尾草屬的研究者並不那麼容易滿足,這些看似相同的植物,是否在內部結構上有細微的不同呢?
當研究團隊仔細檢視該植物的花朵結構時,他們發現了事有蹊蹺,這種鼠尾草的花粉並不像奧地利鼠尾草那樣,被設計成附著在訪花昆蟲的身體側面,而是透過一種更為特殊的機制,使花粉附著在昆蟲的背部。這樣的差異看似相當微小,但其實在鼠尾草屬植物中,這種花粉傳播機制的變化往往彰顯了不同物種間的巨大差異。
鼠尾草的「杠桿」
既然提到了鼠尾草的花粉傳播機制,那麼有必要詳細說明一下這是什麼樣的機制。鼠尾草屬植物特異的雄蕊結構運用了杠桿原理,當昆蟲進入花朵採蜜時,就會推動這個特化的杠桿結構,讓花粉沾附到昆蟲身體上的特定部位,這意味著花粉可能被放置在昆蟲的頭部、背上、甚至腹部或側面。不同鼠尾草的杠桿設計可能因為適應不同昆蟲的行為,形成不同的設計。
在這個故事裡,我們的主角—後來命名為解明鼠尾草的新種—選擇了讓花粉集中沾在於訪花昆蟲的背部,而對比之下,奧地利鼠尾草的花是設計成讓花粉沾附於昆蟲的側面。這種小小的差異,就可能造成物種間的生殖隔離,在長期的持續作用下,這兩個物種的差異將可能持續擴大,走上分家的道路。
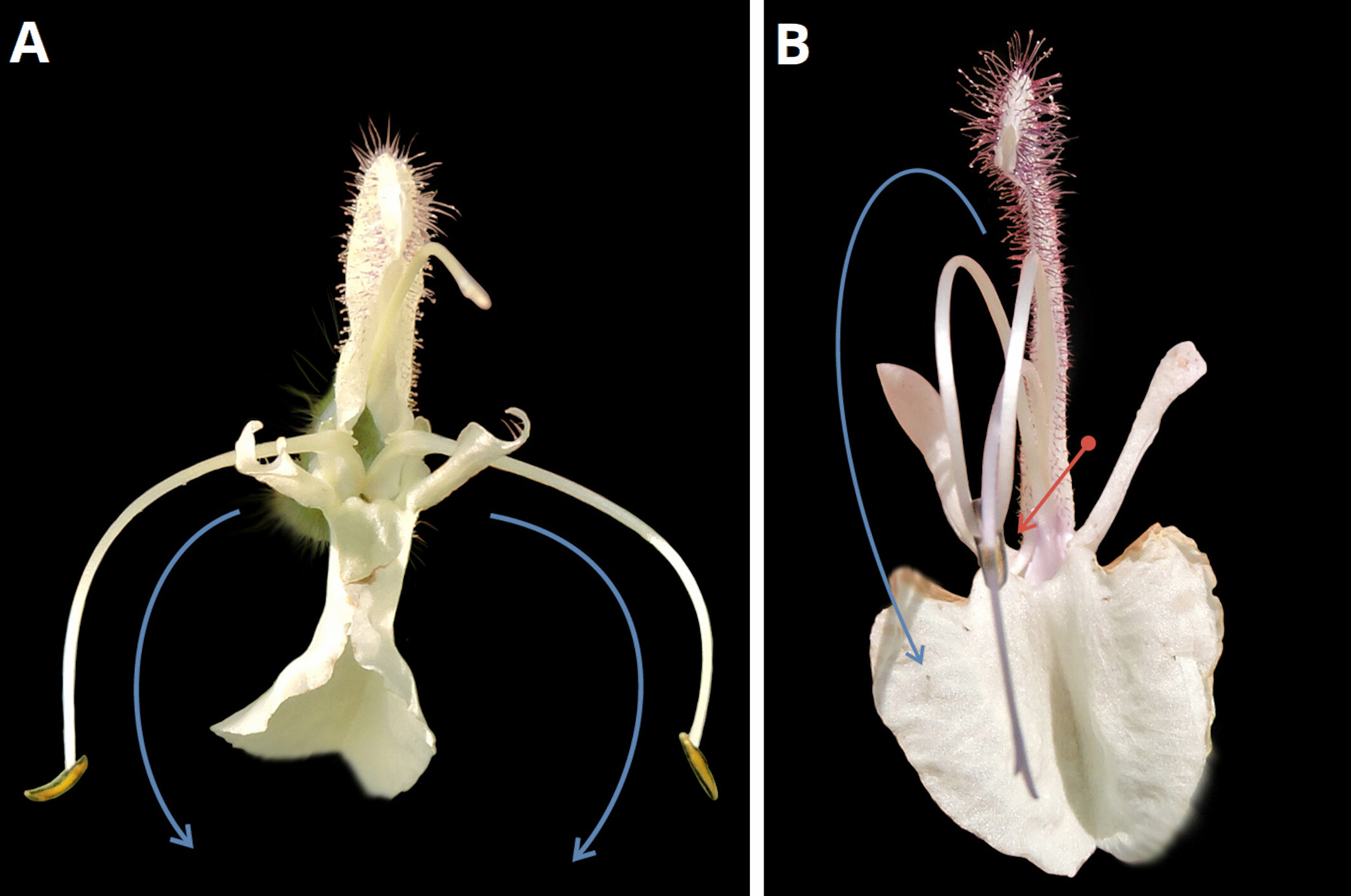
科學破案:形態學、分子證據與地理分布的三重證據
然而,僅光憑花的結構差異或許還不足以確定一個新種,因此研究團隊還加入形態學、基因組分析以及生物地理學的證據,才解開這棵鼠尾草真正的身份。
誠如前述,研究團隊發現該植物的花雖與奧地利鼠尾草類似,但在某些結構仍存在一致而明顯的不同,尤其是在與昆蟲互動的構造上。
想要確認這個鼠尾草究竟是不是一個新物種,必不可少的是分子證據。研究團隊使用了 RADseq 技術,透過分析 DNA 的單核苷酸多態性,構建了謎樣植物與相近種間的親緣關係樹。結果顯示,這種鼠尾草在基因層面上與奧地利鼠尾草明顯分化,兩者無疑屬於不同的物種。
在地理分布上,這兩個物種也是各行其道。奧地利鼠尾草常見於中歐濕潤的森林草原區,而這個新物種則則侷限於較為乾燥的 Pontic 草原 (短草草原地帶),兩者的範圍幾乎沒有重疊,而這種分布上的隔離更加支持了它們是兩個獨立物種的假設。
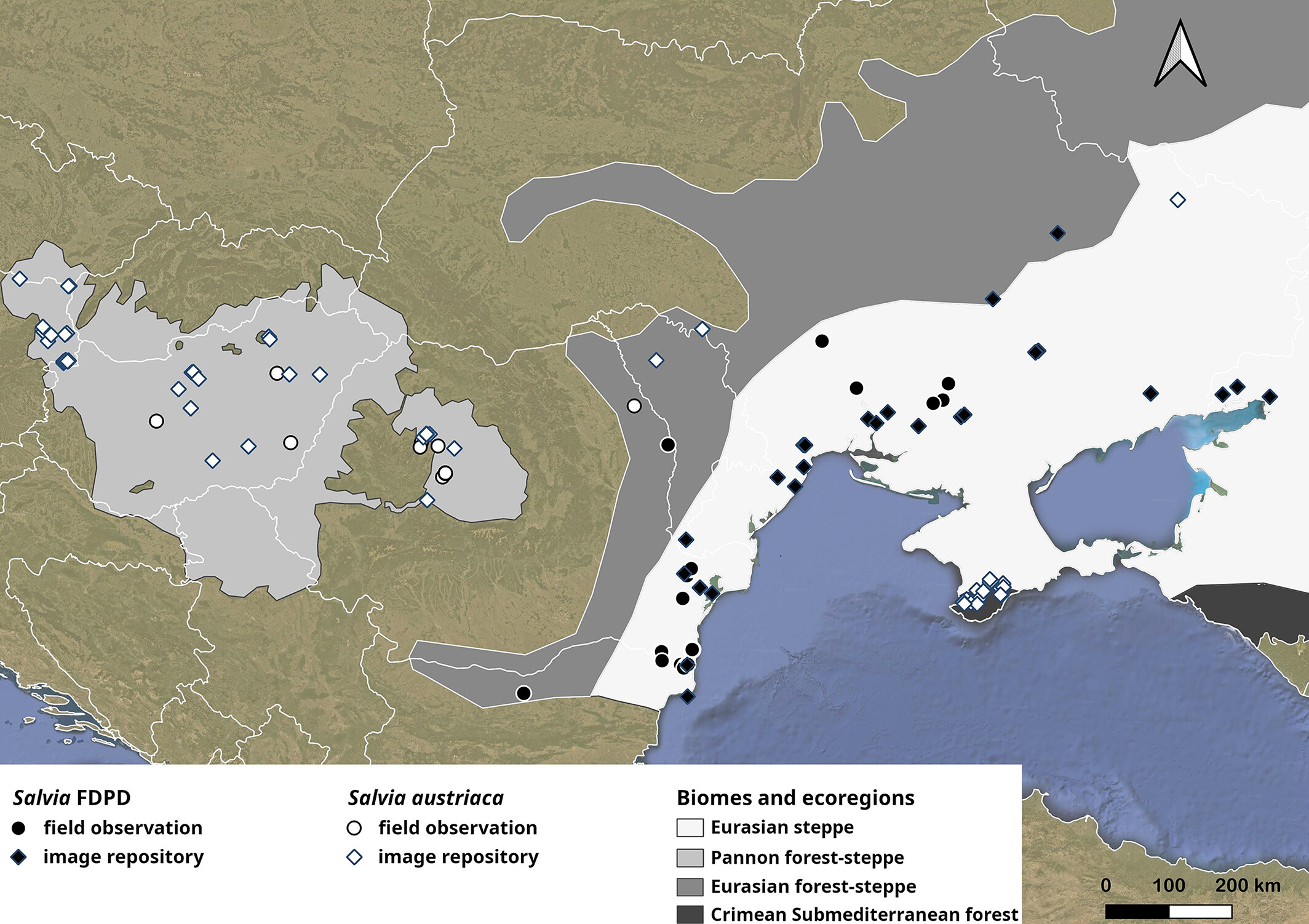
Salvia revelata—一個終於被「解明」的角色
總之,所有的證據都指向新種的誕生,研究團隊將它命名為解明鼠尾草 (Salvia revelata)。這個名字源自拉丁文「revelatus」,意指「被解明的」。這個命名背後是個有趣的隱喻,象徵這個物種長久以來一直在人們的眼前卻未被正視,直到研究者透過細緻的調查,才終於解開這個植物的身份。
有趣的是,這個物種早在 1942 年就已經被人們注意到。當時有人將這種鼠尾草處理為奧地利鼠尾草的一個變種,稱為 Salvia austriaca var. perlanata,這個名稱也被標注在一些標本上。但如今的研究顯示,那些標本實際上應歸入解明鼠尾草,而這個變種名稱現在只能淪為解明鼠尾草的異名。
解明鼠尾草的發現提醒人們,分類學的工作從來不是件容易的事情。即便在熟悉的屬中,仍有可能隱藏許多未被發現的物種。這次的研究還提出,兩個物種的分化可能早在 20 萬年前就已經發生。隨著草原與森林範圍的擴張與收縮,它們逐漸適應不同的棲地,最終各自演化成獨立的物種。
下一步的研究將致力於探討解明鼠尾草的繁殖生物學與保育。此外,這個新的物種是否能與其他鼠尾草基因交流,也是一個有趣的研究方向。總之,分類學只是初始且重要的一步,但分類之外,仍有許多議題等待探索。

結語
看似習以為常的植物,卻可能隱藏無數未被挖掘的故事。解明鼠尾草的故事提醒我們,在自然界中,真正的不同往往藏在細節之中。分類學的醍醐味或許就在於此:越是深究,越能從平凡的表象中挖掘出不凡。而正是這些平凡與不凡,描繪出世界千變萬化的生命多樣性。
Mátis et al. 2022. Hiding in plain sight: Integrative analyses uncover a cryptic Salvia species in Europe. https://doi.org/10.1002/tax.12818

葉齒的暗示
被子植物是地球上最繁盛的植物類群,形態上展現了極大的多樣性。被子植物的許多特徵中,葉片邊緣的葉齒 (teeth),不僅帶來了美感,也是植物分類學和親緣關係研究中的關鍵線索之一。真雙子葉植物(eudicots)約佔所有被子植物種數的四分之三,而葉齒被認為與真雙子葉植物內的某些類群具特殊關聯性。2020年發表的研究就深入探討了葉齒的形態與解剖結構,特別是與葉齒相關的腺體,以了解它們在植物分類、生態和演化 上的意義。
什麼是葉齒?為何重要?
在談到這個研究之前,我們先來了解一下什麼是葉齒。葉齒,顧名思義,是葉片邊緣的突起。其形狀、大小及分布方式千變萬化,有些呈鋸齒狀,有的則如同波浪一般。葉齒在植物上或許不如花朵來得吸引人,但它卻是植物分類上的重要特徵。據統計,葉齒廣泛存在於42個目和173個科中。此外,葉齒被認為是某些據信較為原始的類群(如金粟蘭目 (Chloranthales)、流蘇子目 (Crossosomatales)、或第倫桃目 (Dilleniales)等)的共同特徵,這種特殊的關係,使得它可能在探究被子植物的演化上扮演重要角色。
為了探究這種關係,Rios等學者在巴西的在巴西的採集了47種真雙子葉植物的葉片樣本, 這些葉片包括了10個目、17個科和33屬植物。採取的葉片經過固定、包埋及切片等處理後,利用顯微鏡觀察,並根據前人的研究,把這些葉片的葉齒區分為各種類型。

葉齒形態的多樣性與演化模式
研究結果發現,葉齒形態的多樣性在真雙子葉植物的不同演化支中展現出令人驚嘆的差異。例如,在薔薇類植物(rosids)中,觀察到最多樣的葉齒形態,共包含六種葉齒類型;而在菊類植物(asterids)中,僅發現火把樹型和山茶型兩種葉齒類型,其中火把樹型是最常見的類型。此外,某些獨特的葉齒形態(如刺型和蕁麻型)則僅出現在特定物種中,如冬青屬(Ilex)與瑪雅果屬(Brosimum)植物。更令人感興趣的是,研究顯示某些葉齒形態與植物間的親緣關係密切相關,例如火把樹型葉齒被推測是從薔薇型葉齒衍生而來。這樣的形態在不同親緣支系中可能多次獨立演化,暗示葉齒作為一種適應性結構,在植物的早期演化中就已存在,並且透過趨同演化的模式在不同演化支中反覆出現,幫助植物適應多變的生態環境並發揮多種功能。
葉齒的分類與腺體的關聯
對於葉齒的分類,過去學者曾提出多種方法,最具代表性的是Hickey與Wolfe於1975年提出的15種葉齒類型,包括金粟蘭型 (chloranthoid)、懸鈴木型 (platanoid)、刺型 (spinose)、薔薇型 (rosoid) 等。這些分類不僅描述了葉齒的形態,還強調了它們內部的解剖構造,特別是腺體的存在。腺體是指一類具有分泌功能的構造,這些構造能夠分泌某些特定的物質,來協助植物達到例如防禦或是吸引授粉者等目的,我們熟知的蜜腺 (nectary) 即為一種腺體。
研究發現,腺體與葉齒的關聯性是葉齒分類中至關重要的一環。在這項針對47種真雙子葉植物的研究中,共發現了8種葉齒形態,分別為:秋海棠型 (begonioid)、火把樹型 (cunonioid)、第倫桃型 (dillenioid)、錦葵型 (malvoid)、刺型、山茶型 (theoid)、菫菜型 (violoid) 和蕁麻型 (urticoid)。其中,某些類型的葉齒與腺體密切相關,例如火把樹型中的泌水孔 (hydathodes)、刺型和山茶型中的指狀腺體 (colleters),以及秋海棠型中的花外蜜腺 (extrafloral nectaries, EFNs)。
葉齒中的腺體
研究確認了三種和葉齒相關的腺體類型:
- 泌水孔: 泌水孔是一種特殊的分泌結構,通常用於排出多餘的水分。它們在葉子上的分布具有極高的物種特異性,例如菊科 (Asteraceae)、桔梗科 (Campanulaceae) 或第倫桃科 (Dilleniaceae) 中。泌水孔通常出現在火把樹型或第倫桃型葉齒中。泌水孔常位於葉齒的頂端、表皮具有氣孔,主要集中在腺體的頂端。研究中發現泌水孔的大小和形狀因物種和葉片的發育階段而異。
- 指狀腺體: 指狀腺體是一種分泌粘液的小型結構,主要由單層分泌表皮和內部的薄壁細胞組成。這種腺體會分泌黏液或不溶於水的樹脂,保護幼嫩的器官和分生組織免受脫水和病原體的侵害,在保濕和防禦草食動物和寄生蟲方面發揮作用。指狀腺體的存在與刺型和山茶型這兩種葉齒類型密切相關,特別是在某些黃褥花目 (Malpighiales) 植物。
- 花外蜜腺: 花外蜜腺是一類常見於植物葉片甚至莖部的結構,用於吸引捕食性昆蟲作為防禦機制。它們常位於秋海棠型和山茶型葉齒的上部,為一種小型的球狀結構。
這些腺體都具有特殊的生態功能,也因此,與其相關的葉齒類型,是否反映了植物的生態習性或是棲地特性,仍然是耐人尋味的問題。
葉齒在分類學與親緣研究中的價值
本研究中,研究團隊特別強調葉齒在分類上的應用,例如許多科與目具其特定的葉齒與腺體的組合,而這些特徵若能被充分運用,將有助於解決困難的分類學問題。此外,葉齒形態的演變模式,表明它可能成為親緣研究的重要特徵。
研究顯示,在薔薇類植物(rosids)中,不同葉齒的類型具有一定的穩定性,這對於了解該類群內部的親緣關係可能非常有幫助。此外,葉齒是否帶有腺體及腺體的種類,在薔薇類植物的不同科之間表現出特定的模式。這暗示了葉齒和腺體的組合可能在薔薇類植物的演化過程中,受到某些特定的環境或生物因素的影響而產生適應性變化。
結論
葉齒是真雙子葉植物中一種雖然常見但長期被低估的構造,其形態和解剖特徵蘊含著豐富的訊息。本研究不僅發現葉齒的形態多樣性,還說明了葉齒與腺體的組合在分類中的潛在應用角色。更重要的是,這項研究打開了一扇探索葉齒功能與生態意義的大門,並促使人們重新審視葉齒在植物科學中的地位。未來,科學家可進一步探討葉齒在植物適應環境、吸引益蟲或防禦捕食中的功能,以及葉齒的演化與植物親緣的關係。
Rios et al. 2020. Leaf teeth in eudicots: what can anatomy elucidate? https://doi.org/10.1093/botlinnean/boaa028

蝴蝶翅膀不只漂亮,還很有用!
這幾年,新聞報導裡時不時可以看到氣候變遷對蜜蜂生態的影響,各位可能多少有聽說蜜蜂對農作物的重要性,與其在農業生產中扮演的不可或缺的角色。牠們高效率的傳粉能力,不僅讓農田的作物結實累累,也維繫著天然生態系中的植物繁殖。但能夠為植物傳粉的動物可不只有蜜蜂,蝴蝶、鳥類甚至蝙蝠都可能是植物的合作夥伴,幫助完成花粉的傳遞。然而,你是否想過,「蝴蝶翅膀」也能成為植物傳宗接代的重要橋梁?這聽起來有點不可思議,不過在南非的一些植物中,這卻是一種既特別又有效的授粉方式。近期針對火球花 (Scadoxus multiflorus) 的研究闡明了這種植物與蝴蝶之間的互動,也讓我們重新認識這些演化下的天擇適應。
要認識這個蝴蝶翅膀傳粉的現象,我們得先簡單了解什麼是動物訪花與授粉效率。這個世界上會拜訪花的動物很多,然而並不是所有前來的動物都能有效地授粉,有些動物可能只是來採蜜卻沒有碰到雄蕊或雌蕊。所以對植物來說,讓訪花者「精準」地帶走並傳遞花粉是一門需要長期演化的技術。火球花就是個很好的例子,研究指出,這種植物並不是像其他植物利用訪花昆蟲的腳或腹部,而是專注於蝴蝶的翅膀來傳宗接代。
為蝴蝶翅膀量身打造的花朵設計
火球花 (Scadoxus multiflorus ) 是石蒜科的一種草本植物,主要分布於南非,有著紅色、鮮豔的花,吸引著以大型蝴蝶為主的傳粉者,臺灣也有許多人在家中栽培這種植物。然而,這些蝴蝶並不像植物常見的傳粉者那樣,透過身體直接接觸花葯和柱頭來傳遞花粉。相反地,火球花將花粉「安放」在蝴蝶翅膀的腹側,一個對蝴蝶來說似乎毫無影響的位置。

研究發現,火球花的兩個亞種各自有不同的主要傳粉者:S. multiflorus subsp. katherinae 主要依賴鳳蝶科Papilio dardanus cenea 的雄性,而另一亞種 S. multiflorus subsp. multiflorus 則依賴鳳蝶與粉蝶的合作。這種傳粉行為的模式相當專一,這些植物幾乎完全依賴這些特定蝴蝶完成授粉,因為火球花高度自交不親和,無法透過自花授粉產生種子。
植物如何確保蝴蝶翅膀能有效黏上花粉並帶到下一朵花?一切都要歸功於火球花特別的花朵結構。研究發現,火球花的花藥與柱頭分散排列,並突顯於花朵表面,這種「開放刷狀」結構使得當蝴蝶在花朵間飛翔吸蜜時,翅膀會不可避免地接觸花朵關鍵部位,從翅膀帶走花粉,又將花粉放到下一株植物的柱頭上。
蝴蝶翅膀授粉的多重起源
火球花或許是蝴蝶翅膀授粉的代表性植物,但研究顯示,這種模式並非僅此一例。南非至少有9種石蒜科植物可能存在類似的授粉方式。像是知名的君子蘭 (Clivia miniata)、Brunsvigia marginata 和 Cyrtanthus elatus 等,都具有將花粉黏在蝴蝶翅膀的授粉模式。雖然這些植物的花葯位置、花形或花色有所不同,但牠們的核心授粉策略卻都一致地與蝴蝶翅膀有關,很明顯這是一種由天擇塑造的趨同演化。
這些石蒜科植物的花朵各自表現出一定程度的特化,例如君子蘭的花較為「碗狀」,即花葯集中於花瓣邊緣,而非分散排列。當蝴蝶探入花瓣內部吸蜜時,其翅膀自然會沾染上花粉,和火球花的「開放刷狀」設計大相逕庭,但目的卻如出一轍。
接下來就不能不提另一個值得注意的現象,研究指出,這些蝴蝶翅膀授粉的機制可能多次、獨立地起源於石蒜科的不同植物中,這意味著即使分布區域重疊,各植物都找到了自己的「蝴蝶合作方案」,演化出適合自身需求的傳播策略。
城市化與蝴蝶的消失
任何一個生態系的合作關係都建立在傳粉者族群的穩定性上。然而,隨著人類活動擴大,傳粉者族群已成為生態系中最為脆弱的一環。研究發現,在某些都市化區域,由於蝴蝶數量匱乏,Scadoxus multiflorus subsp. katherinae 的結果率顯著下降。不僅如此,科學家計算得出,該地點的花粉轉移效率僅為 1.47%,這意味著絕大多數的花粉並未成功達到宿主植物的柱頭上。
蝴蝶的數量下降可能是多種環境壓力的共伴結果,包括棲地喪失、大量使用農藥、氣候變遷等。雖然大多數植物學研究聚焦於植物本身的適應性與生態需求,但這裡也再次提醒我們,傳粉者的保育工作直接影響植物繁殖與生態系統的平衡,就像文章開頭提到蜜蜂的重要性一樣。
結語
從火球花與蝴蝶的互動,我們可以看到天擇如何塑造不同物種間的合作,也讓我們看見當生態網路的一環中斷時可能產生的連鎖反應。不論是蝴蝶翅膀是否能成功帶走花粉,還是其族群數量能否穩定維持,這些看似細微的互動,都支撐著一個物種的繁殖生存之路。
植物本身無法移動或選擇自己的伴侶,但透過長久的適應與演化,它們成為生態系中不可或缺的核心角色。而蝴蝶翅膀上的花粉,從一朵火球花傳到另一朵火球花,不僅讓人驚嘆於自然界的巧妙設計,更提醒著我們,這場時刻進行的傳粉生態舞台,是人類依存的環境當中不起眼但需要保護的一部分。
Butler, H.C. and S.D. Johnson 2020. Butterfly-wing pollination in Scadoxus and other South African Amaryllidaceae. https://doi.org/10.1093/botlinnean/boaa016

小心!別吃到有毒的花蜜
2014年6月,福建省西北部的泰寧縣發生了一件駭人聽聞的事件,有19名村民在食用了野生蜂蜜後發生中毒。這些村民被緊急送往醫院救治,幸運的是有16名村民經搶救後脫險,但仍然有3名村民不治而亡。有人推測,這些村民食用的蜂蜜中可能含有劇毒植物雷公藤 (Tripterygium wilfordii, 衛矛科) 的花蜜,這才導致中毒。事件本身已經引人側目,但其中更令人好奇的是——一直以來都帶著甜美、無害印象的花蜜,為什麼會有毒?
在日常生活中,花蜜給人的印象通常是甜美、無害且富有吸引力,很多人應該都有過品嚐朱槿 (Hibiscus rosasinensis) 或仙丹花裡的甜甜花蜜的經驗。而從生態學的角度來看,花蜜就是植物提供作為「報酬」的甜味分泌物,用來吸引並回饋幫助授粉的動物。基於這樣的理解,我們似乎很難將花蜜與「有毒」聯想在一起。然而,這並不是一個單獨的現象,而是自然界真實存在的一部分。所以,今天就讓我們以此為切入點,一起來探討有毒花蜜背後的生態學意義吧!
花蜜是怎麼形成的?
花蜜 (nectar) 是開花植物透過一種稱為蜜腺 (nectary) 的特殊構造分泌出的含糖混合物。不過有趣的是,蜜腺不僅分布在花朵,以吸引授粉者,有些蜂蜜腺還可能生長在葉柄等其他部位,稱為花外蜜腺 (extrafloral nectary)。像山櫻花 (Prunus campanulata) 葉柄上的腺體就是典型例子。不過今天的討論主要聚焦在花朵上的蜜腺。
花蜜的成分裡,90%左右為糖,剩下約一成的成分包括水分、脂質、氨基酸、抗氧化物、礦物質及次級代謝物。不同植物的花蜜成分差異極大,糖與次級成分比例上的細微變化,造就了不同植物花蜜的特殊風味,也讓我們能從蜂蜜的品嚐中略知其來源。而在這些成分中,致毒的元素大多是植物自身的次級代謝物。這些次級代謝物遍布植物體內,而後經由自然的運輸過程進入花蜜。
有毒花蜜常見嗎?
有毒花蜜,顧名思義,指的是含有能對訪花動物產生驅避或毒害作用成分的花蜜。由於有毒的次級代謝物是植物生存策略的一部分,因此它們進入花蜜中並不讓人意外。有研究列出了至少30個不同植物科含有這樣的有毒花蜜,較為知名的例子包括杜鵑花科 (Ericaceae)、茄科 (Solanaceae)、百合科 (Liliaceae) 等植物。
其中最著名的可能是杜鵑花 (Rhododendron) 的花蜜,其含有一種名為梫木毒素 (Grayanotoxin) 的化合物。攝取過量杜鵑花蜜中的梫木毒素可導致狂蜜症,其症狀包括呼吸困難、低血壓及心律不整。然而,若劑量極少,梫木毒素卻可能引發迷幻效果,因此少數人甚至刻意服用,以追求這樣的感受。

為什麼植物會產生有毒的花蜜?
從上述例子可以看出,有毒花蜜十分普遍,但它的出現絕非偶然,而可能有深層的生態意義。針對這一點,學者們提出了一些有趣的假說:
- 授粉者專一性假說:這個假說認為,有毒花蜜能有效鼓勵特定授粉者的專一性,並排除不必要的訪花者。例如某些授粉者能適應花蜜中的毒素,反倒可以更專注地幫助植物傳粉,進一步提高效率。
- 花蜜竊盜者阻嚇假說:此假說主張,有毒花蜜的毒成分可以防禦非目標訪花者,例如螞蟻或其他竊取花蜜卻不幫助授粉的動物。以梓樹 (Catalpa speciosa) 為例,其花蜜含有環烯醚萜苷 (iridoid glycosides),可有效驅趕螞蟻及弄蝶,但對蜜蜂等授粉者卻影響不大。
- 「醉酒」授粉者假說:某些植物或許不是要驅逐訪花者,而是希望藉由毒素改變訪花者的行為。以火燒蘭屬 (Epipactis) 花蜜為例,其含有乙醇成分,可能導致訪花的蜜蜂稍顯遲鈍,減少整理身體的次數,因而增加花粉的有效傳遞。
- 抗菌假說:這一假說側重花蜜自身的保存,認為毒素中的某些化合物可能具有抗菌功能,保護花蜜不被微生物降解,進而長時間維持吸引力。
- 多效性假說:最後,此假說認為,有毒花蜜或許只是植物產生次級代謝物作為抗草食性防禦的一個副產品,並不是有意為之。這些化合物透過韌皮部擴散到花蜜中,結果形成一種「附帶效應」。
雖然看似有許多合理的假說,但目前的實證研究仍有限,多數重點僅在於描述有毒花蜜的存在以及其可能功能,缺乏直接的實驗數據來證實這些假說的正確性。例如,有毒花蜜是否真的能帶來生態學優勢?它的成分變化是否具有遺傳基礎?這種策略究竟是自然選汰的結果,還是一種無害的副作用?這些問題仍然有待探索。
有毒的花蜜,雖然與我們對甜美無害的印象相悖,但卻展現了植物在大自然中精妙的生存智慧。透過漫長的演化,即使無法像動物那樣自由移動,植物依然發展出許多驚人的策略,而花蜜中的毒性正是其中之一。然而,目前多數關於有毒花蜜的研究仍停留在理論層面,缺乏足夠的實驗證據來驗證其真實的生態功能,許多問題依然懸而未解。不過,這也正是科學迷人的地方,每個未解之謎都在等待更多人一同參與探索。生態研究的進步不僅仰賴技術與資源,更需要新一代研究者的加入,無論是熱愛自然的學生,還是對科學充滿好奇的每個人,你的視角與努力都可能成為解開這些謎團的重要力量。面對如此豐富且未知的自然界,或許下一頁的答案,就在等待你來翻開。
Adler, L.S. 2000. The ecological significance of toxic nectar. https://doi.org/10.1034/j.1600-0706.2000.910301.x

2024臺灣新植物
時光飛逝,又過了一年。老實說,這個單元已經連載了好幾年了,但每年臺灣仍然持續有許多新的植物被發現,其中相當高的比例是新種或新紀錄種。這些新發現的原生植物,清楚地反映了我們對臺灣的探索與認識,仍有許多尚未被揭開的篇章可供追尋。臺灣的許多角落至今仍未被人們充分認識,許多地區也尚未被植物學者或愛好者深入探訪,這些地方有可能潛藏著許多新的植物,等待我們去發掘。
然而,近幾年也發現了不少新的原生植物,其棲地並不一定處在人跡罕至的深山或偏僻之地,這表明,即使是在我們熟悉的環境中,或許依然蘊藏著新的驚喜與可能性。同時,許多植物族群的分類地位也可能需要再深入檢討與更新。這些發現與研究顯示,無論是既有的認知還是全新的探索,我們對這片土地的理解仍有很大的成長與發展空間。而那些被報導的外來種,代表著臺灣與世界各國的交流下,有意或無意引進的新住民,這些植物由於不是臺灣的原生物種,或許代表著潛在的生態危機,同樣也值得我們的研究與注意。
新種
- 卑南懸鉤子Rubus puyumaensis J.Y. Huang, P.H. Chen & A.C. Chung
https://bdj.pensoft.net/article/115431/ - 中央尖山艾 Artemisia tyuosenicola S.W. Chung, W.J. Huang & T.C. Hsu
tai.2024.69.372
種小名是源自於古老的標本中對中央尖山的地名拼字 - 玉蜘蛛蘭 Taeniophyllum lishanianum T.P. Lin & G.X. Zhu
- 塔山蜘蛛蘭Taeniophyllum tumulusum T.P. Lin & G.X. Zhu
tai.2024.69.302 - 大雪山蜘蛛蘭 Taeniophyllum daxueshanensis T.P. Lin
- 大扁根蜘蛛蘭 T. taiwanensis T.P. Lin
- 輻射糙莖皿柱蘭 Lecanorchis purpurea var. actinomorpha T.P. Lin
https://taiwania.ntu.edu.tw/pdf/tai.2024.69.380.pdf - 凌雲巖油點草Tricyrtis lingyunyanensis Y.Huan Chen & T.H.Huang
https://phytotaxa.mapress.com/pt/article/view/phytotaxa.655.3.2
很像毛果油點草 (T. lasiocarpa) XD - 勝任鐵線蓮C. parviloba Gard. ex Champ. subsp. shengzhenii Y.H. Cheng & T.Y.A. Yang
- 彭氏鐵線蓮C. pengii Y.H. Cheng & T.Y.A. Yang
https://phytotaxa.mapress.com/pt/article/view/phytotaxa.662.1.2 - 阿禮當歸Angelica aliensis H.H. Chen & J. C. Wang
https://as-botanicalstudies.springeropen.com/articles/10.1186/s40529-023-00407-7
終於發表的新種,長在很神秘的霧台深山。
新紀錄種
- 瓦氏挖耳草 Utricularia warburgii K.I. Goebel
https://taiwania.ntu.edu.tw/abstract/2036 - 毛當歸 Angelica pubescens Maxim.
https://as-botanicalstudies.springeropen.com/articles/10.1186/s40529-023-00407-7
這篇文章同時也把南湖當歸A. morrisonicola var. nanhutashanensis Liu, Chao & Chuang處理為玉山當歸A. morrisonicola 的同物異名。
新馴化種
- 立性酢漿草Oxalis barrelieri L.
- 砂莧Allmania nodiflora (L.) R. Br.
https://www.tbri.gov.tw/view.php?theme=publications&subtheme=biodiversity&id=80&font-size=m - 寬葉雀稗 Paspalum mandiocalum Trin
- 皺稃雀稗 Paspalum plicatulum Michx.
- 絨葉雀稗 Paspalum virletii E. Fourn.
生物多樣性研究所 - 毛耳節藤 Camonea pilosa (Houttuyn) A.R.Simões & Staples
國立臺灣博物館學刊77-1期 - 麻葉鼬瓣花 Galeopsis tetrahit L.
https://www.tbri.gov.tw/view.php?theme=publications&subtheme=conservation&id=147
註:直至截稿日,林業研究季刊的第46卷尚未上線,這個期刊過往報導過相當多的新類群,待日後更新再將相關資料補上。

學名長得太像怎麼辦?
所有修過植物分類學的學生,大概都曾經經歷過背學名的地獄,雖然說現在的教育傾向理解而不推崇背誦,不過一些常見植物的學名還是免不了要記下來。不過植物分類學可是一門發展了超過三個世紀的學問,歷來發表過的學名多不勝數,這麼多發表過的學名裡面,有沒有剛好拼字完全一樣的呢?答案是當然的,而且其實還不少,這類名稱又稱為同名 (homonym),根據命名法規第53條款的規定,這種同名的狀況中,比較晚發表的名稱是不合法的,必須提出一個替換名 (replacement name) 來代替使用。
不過當遇到的是長得很像的學名,這種狀況就有點討厭了,因為理論上只要拼字不是完全一樣的學名,都應該被視為不同的名稱,所以只要發表過程合於法規,就會是一個合法的學名。這種案例非常得多,例如說Prunus armeniaca和P. americana或是P. caroliniana和P. carolinae等,儘管這些名稱可能源自於不同的語源或是意義,然而長得很像還是讓人非常容易搞混。以臺灣的植物為例的話,大概就是一堆以臺灣 (Formosa) 為名的種小名 (species epithet) 最容易讓人搞混了。
有鑑於此,法規中的第53條款中,說明這類過於相似的名稱,如果已經造成應用上的混淆時,那麼可以把較晚發表的名稱視為同名處理。不過由於這種相似性是否會造成混淆實在很難認定,所以法規的53.4條也說明,當有這類狀況的疑慮時,可以把案例提交給法規的總委員會 (general committee) 審查,如過經投票通過,那就會做成約束性的決議,並且記錄在附錄II中。
以下就以一個臺灣的案例來說明實際執行起來的流程和狀況。臺灣胡頹子 (Elaeagnus formosana Nakai) 和蓬萊胡頹子 (E. formosensis Hatus.) 是兩種臺灣的特有種胡頹子,很明顯,兩個種類的種小名都是源自於臺灣(Formosa)這一名稱,且語法上都屬於形容詞。但一個是以形容物種來源的形式(formosensis,意為「來自臺灣」),而另一個則是直接以「福爾摩沙」相關的形容詞形式(formosana,意為「臺灣的」)來表述,所以被認為是容易混淆的學名。黃增泉認為這種狀況應該可以根據法規的規定,將晚發表的E. formosensis視為E. formosana的同名,因此在2002年提出了一個E. formosensis的替換名E. ohashii,即大橋胡頹子,語源是紀念日本東北大學的大橋廣好 (H. Ohashi) 教授。

不過究竟這個名字的替換是不是有必要的呢?中國學者鄧雲飛在2007年向IAPT的總委員會,針對上述兩個名稱的混淆問題提出申請。鄧雲飛在提出的申請說明中表示:
最近,我注意到 Elaeagnus formosensis的命名問題。在胡頹子屬 (Elaeagnus ) 中,有兩個源自「Formosa」的名稱,即 E. formosana Nakai和 E. formosensis Hatus.。Huang依據第53.3條規定,將這兩個名稱視為同名,並提出以新名稱 E. ohashii Huang 取代 E. formosensis的名稱。
然而,我認為這兩個名稱不應作為「同名」處理,因為它們代表「Formosa」的兩種不同型式。這與第53條範例第9例中 Lysimachia hemsleyana Oliv.和 L. hemsleyi Franch. 的情況類似,既然上述兩者不被視為容易混淆的名稱,那麼本案例中的E. formosensis和E. formosana也不應該。因此,E. ohashii 是一個多餘的名稱 (superfluous name)。我的理解正確嗎?
維管束植物命名委員會 (Nomenclature Committee for Vascular Plants) 針對這件申請投票,投票結果有8票贊成、10票反對的狀況下,認為這兩個名字不致造成混淆,根據投票結果,E. ohashii應該作為一個多餘名看待。而總委員會在2019年的報告,也再次的確認了這個結果。目前,這個案例已經列在法規的附錄七當中。
這個案例展現了植物命名法規在處理相似名稱時的審慎與複雜性,並且彰顯了法規的核心理念,即保持學名使用的穩定性。透過這個案例,可以看出命名法規不僅是植物學家在學術研究中的工具,更是在維護命名秩序與促進國際合作上的重要基石。隨著更多新物種的發現與分類,命名法規也會持續演變,但其核心精神——穩定性與精確性——將始終是分類學的指引。
Brummitt, R.K. 2007. Report of the Nomenclature Committee for Vascular Plants: 58. https://doi.org/10.1002/tax.562032
Barrie, F.R. 2011. Report of the General Committee: 11. https://onlinelibrary.wiley.com/doi/pdf/10.1002/tax.604026