
看走眼的誤會,從酒瓶蘭亞科到鈴蘭亞科
APG系統的主流化已經不是新聞,很多我們熟悉的科也迎來重大變動,其中比較為人熟知的包括玄參科 (Scorphulariaceae)、車前科 (Plantaginaceae)、楊柳科 (Salicaceae)、錦葵科 (Malvaceae) 以及百合科 (Liliaceae) 等。其中百合科在細分之後,反而產生了另一個大科—天門冬科 (Asparagaceae),根據APG系統第四版的分類架構,這個科可以分成7個亞科,而其中之一就是酒瓶蘭亞科 (Nolinoideae)。
說起酒瓶蘭亞科可能大家不太熟悉,不過這個亞科包括許多日常生活中常見的植物,例如巴西鐵樹 (Dracaena fragrans)、黃精 (Polygonatum)、蜘蛛抱蛋 (Aspidistra)。不過這個亞科的學名最近卻被認為是有問題的,到底是出了什麼事呢?

長期以來,酒瓶蘭亞科的學名Nolinoideae一直被認為是由Burnett在1835年發表的名稱,但是最近有研究人員發現,當時Burnett在文獻中紀錄的其實是茄科的Nolana,而非酒瓶蘭屬Nolina,從Burnett提供的形態敘述看來,他指的的確是茄科植物,因此並非筆誤。所以當時Burnett成立的亞科是Nolanoideae,而不是Nolinoideae。
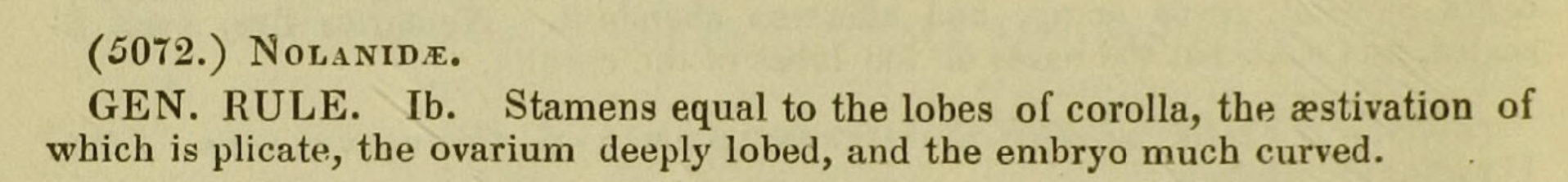
Fischer在2015年也發現了這個問題,所以他以酒瓶蘭屬為模式,正式的提出酒瓶蘭亞科 (Nolinoideae) 的合法名稱。然而,法規的常識告訴我們,最早發表的合法學名才具有優先權。近期的研究指出,這個亞科最早的合法名稱,應該是Herbert在1837年發表的Convallarioideae,這個亞科的模式是鈴蘭屬 (Convallaria),也因此過去所稱的酒瓶蘭亞科,其實應該是鈴蘭亞科。
Tanaka & Nguyen 2022. Nolinoideae (Asparagaceae) in APG III needs replacing with Convallarioideae. https://doi.org/10.11646/phytotaxa.583.3.9

馬祖的換錦花究竟是誰?
每年8月,有許多愛花人趨之若鶩的前往國之北疆的東引島上,這些人的目的不為別的,正是那島上盛開的石蒜花—換錦花。換錦花屬於石蒜科 (Amaryllidaceae),這個科有不少耳熟能詳的觀賞植物,例如孤挺花 (Hippeastrum)、石蒜 (Lycoris)、文殊蘭 (Crinum)、蜘蛛百合 (Hymenocallis)、水仙 (Narcissus) 等,就連經常供食用的蔥屬植物 (Allium) 也是這個科的成員之一。

說到原生的石蒜屬植物,臺灣島上可見的僅有金花石蒜 (L. aurea) 一種,生長於中北部的岩場,每年約在9-10月開花。而另外兩種紅花石蒜 (L. radiata) 和換錦花 (L. insularis, 海濱石蒜) 則僅見於馬祖。其中又以換錦花的數量最少,僅見於東引和西引兩島上。過去多認為換錦花與產於中國的L. sprengeri同種,不過去年的一個研究卻指出事實並非如此。
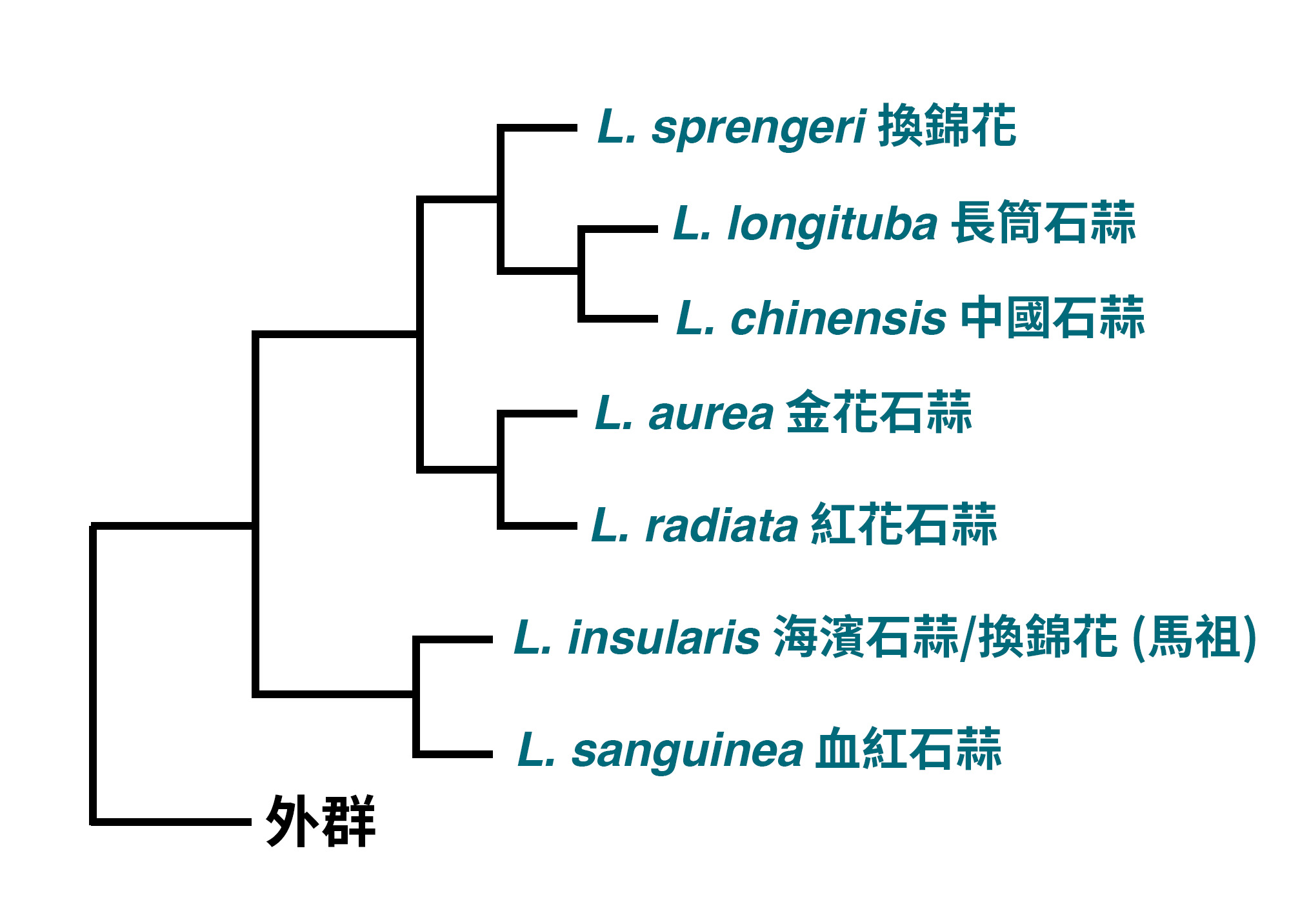
Zhang等人指出,過去認為L. sprengeri廣泛的分布於中國的安徽、江蘇、浙江及福建各省,不過從分布上看來,明顯的可以發現區分為安徽內陸至湖北的內陸地區,以及江蘇至福建等沿海地區的兩大群體。研究人員在檢視了兩大群體的形態後,發現沿海族群的花冠筒較內陸者更長,不過僅有這樣的特徵似乎很難說明這個群體是不同的種類。於是研究人員又分別研究了這兩個群體的親緣關係,根據葉綠體基因組重建的親緣關係樹,兩個群體各自形成支持度高的分支,兩者的親緣關係亦不接近,這樣的結果加上先前的形態差異,說明兩者確實為不同的種類,因此沿海地區的群體被發表為一新種,即Lycoris insularis,作者提供的中文名稱為海濱石蒜,而根據報告中的分布地圖看來,這個新種的分布也包括了馬祖地區的族群,因此馬祖的換錦花,實則為這個新種。

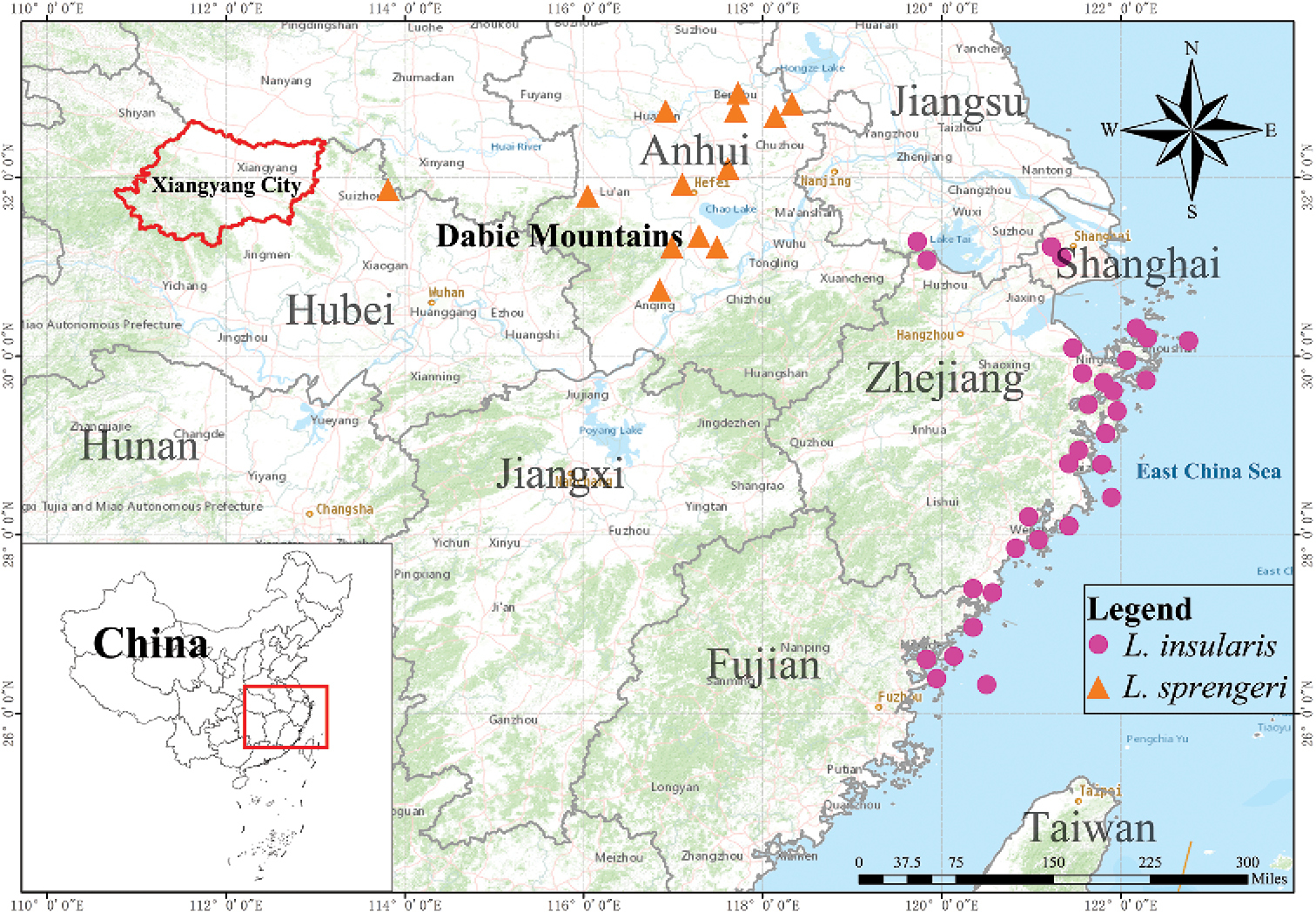
至於那些換錦花與紅花石蒜雜交的個體名稱,則不受到這樣的學名變更的影響 (這些雜交個體其實也還沒有被正式的命名)。
Zhang et al. 2022. Lycoris insularis (Amaryllidaceae), a new species from eastern China revealed by morphological and molecular evidence. https://phytokeys.pensoft.net/article/90720/

多久了、我都沒變,愛你這回事、整整六年—愛不到就直接長過去之花粉管淋浴
野牡丹科 (Melastomataceae) 是被子植物中一個很大的科,約有175屬、1500-1600種植物,廣泛分布在全世界的熱帶至亞熱帶地區。本科植物的外型也很多變,包括一年生至多年生草本、灌木、小喬木等。不過談到野牡丹科,最為人所知的大概是其特殊的授粉模式,本科植物的雄蕊花葯通常孔裂,這種方式其實不利於花粉的散播,必須仰賴蜂類昆蟲停留在雄蕊上時振動翅膀,以這樣的振動頻率把花粉散出,再藉由這些蜂把花粉帶到另一朵花的柱頭上。這種授粉模式被稱之為震動授粉 (buzz pollination),除了野牡丹科外,也在其他多種植物中發現。

這種授粉方式的好處是可以降低自花授粉 (autogamy) 的機率,避免其帶來的不良影響,不過缺點也顯而易見,當你長在一個蜂類稀少的地方,那你的花成功授粉的機率就大大降低。到時候別說自交造成的不良影響,連能不能延續香火都成了問題,很多植物在這種授粉昆蟲稀缺的情況下都發展出延遲自交 (delay selfing) 的機制,讓這些花可以藉由自交產生種子,得到延續後代的保障。所謂的延遲自交指的是這些花在剛開放時為傾向異交,然而在經過一段時間後若仍然沒有授粉者拜訪,那麼這些花就會啟動自花授粉的機制,利用自花授粉的方式產生種子。延遲在這裡形容的應該是相對於閉鎖花等從一開始便是自花授粉的花而言,是直到開放後期才具有自花授粉的現象。然而野牡丹科的花朵構造特殊,雌雄蕊間的距離較長,加上花朵發育過程中兩者無法接觸,因此過去許多研究都認為野牡丹科基本上沒有自花授粉的機制,但在科學家發現兩種巴西產野牡丹科植物的授粉方式後,讓人大大改觀。
華貴草屬 (Bertolonia) 是一群包括約35種、南美洲特有的野牡丹科植物,本屬多為森林底層低矮的草本地被植物,僅產於巴西大西洋側的森林中。花為白色至粉紅色,雄蕊具有典型的孔裂花葯。許多人對這屬植物的印象,大多在於顏色多變且具有絨布質感的葉片 (可能也是中文名稱的由來),因而可見觀葉市場中流通。巴西的熱帶雨林中兩種華貴草屬植物 (B. paranaensis和B. mosenii) 被發現具有延遲自交的現象,不過就像上面說的,野牡丹科的雌蕊和雄蕊的距離之遠,加上特殊的花粉散播機制,要自花授粉的難度可說是比牛郎和織女更難 (好老的梗),不過這兩種植物的雄蕊硬是利用了非常特殊的方式,跨越這不可能的距離。

研究人員發現,這兩種植物的單朵花的開放時間大約持續一天,從每天早上4點開始,到大約6點左右花全部開放,這個時候雌雄蕊彼此還頗有距離不接觸。然而到了下午4:30左右,花朵開始閉合,這個時候雄蕊的花絲逐漸開始移動靠近花柱,直到下午7點左右,花朵完全閉合,而這個時候雄蕊完全靠在花柱上,花葯上的孔直接正對柱頭!不過僅僅這樣還是沒用,前面說過這些花粉沒有震動就休想逃離花葯的控制,只要沒辦法離開花葯,就算雄蕊直接貼在柱頭上還是沒有授粉的可能。不過花粉出不去不打緊,只要能夠把精細胞從花粉送出去就可以,這些花粉就在這個時候開始萌發花粉管 (pollen tube),藉由萌發花粉管的方式離開花葯,讓花粉管直達柱頭,把柱頭密密麻麻的包覆起來,接下來就是植物通常的授粉步驟,精卵結合,胚珠發育為種子。研究者還把這樣的現象稱為花粉管淋浴 (pollen tube shower),可能是覺得那個花粉管從花葯齊發的樣子很像從蓮蓬頭噴射的水柱吧?

除了這種近似恐怖情人的延遲自交機制外,筆者認為這個研究的另一個亮點在於過去許多這類研究著重的是花與授粉者互動的多樣性,但是這個研究卻揭示了:就算沒有授粉者的存在,自交的機制也同樣的神秘而炫目,也同樣值得我們的研究。
Passos et al. 2022 “Pollen tube shower” in Bertolonia (Melastomataceae): a new delayed selfing mechanism in flowers with poricidal anthers. doi.org/10.1093/botlinnean/boab051
註:原始研究文獻上的圖很精彩,充分展現出花粉管淋浴的樣貌,但因版權問題在此不便提供。

我的故鄉在遠方—橫斷臺灣
不知道正在看這篇文章的各位,是不是有玩過幾個月前剛上市的熱門電玩—薩爾達傳說 王國之淚呢?老實說,我從發售日以來沈迷至今,這個遊戲廣大的地圖、各式各樣的地形與生物,主角林克擁有各種不同的能力,可以在這片廣闊的海拉魯大地上四處探險,收集各種素材、解決謎團與戰勝魔王。不過我一邊玩這個遊戲,想到的卻是有關旨价的書稿的主題—臺灣的植物到底從哪裡來?王國之淚中,林克擁有一個特殊能力名為倒轉乾坤,可以把物體根據它過去移動的軌跡,回到特定時空中的位置。現實生活中當然不可能有如此方便的工具,不過植物研究者還是藉著許多殘留至今的線索,探求在漫長的時間裡,植物是如何起源並散布到世界的各個角落,其中當然也包括臺灣。

這種議題在生物研究中屬於一個專有的領域,名曰生物地理學 (biogeography)。這門學科主要探討的是某一類生物如何在地球漫長的歷史中出現,並且逐漸擴散成目前的分布狀況。橫斷臺灣一書要探討的主題就是,一個和臺灣又近又遠的高山國度—橫斷山,以及它與臺灣的連結。說遠,兩者的地理距離長達好幾千公里,但是它其實很近,兩者同樣都是高山國度,也令人意外的擁有相當多共同或是近緣的植物。無論如何,這個高山國度對臺灣的讀者而言是陌生的,或許遙遠的地理距離也是原因之一,不過更多的原因,可能是在於它長期掩蓋在喜馬拉雅山的陰影下。
橫斷山一詞,即使是對研究者來說,仍然是年輕的名詞。這個盤據在中國西南的廣大山地,過去的稱呼包括中國西南山地、東喜馬拉雅山等,長期以來作為附屬品一般的角色存在。然而近年的研究中,逐漸凸現這個區域的重要性。許多被認為起源於喜馬拉雅山的植物,紛紛被證明其實是起源於橫斷山;換言之,喜馬拉雅山更像是這些起源於橫斷山的植物的去向,而不是來處。然而臺灣呢?從日本時代起,就有很多學者注意到臺灣與喜馬拉雅山兩地植物的相似性,相隔甚遠的兩個地區竟然有類似的植物,令當時的學者大感驚奇,然而當時雖然發現了這個現象,但沒有提出造成這種現象的假說。近來的研究說明,其實這個和臺灣有關的地區並不是喜馬拉雅,而是橫斷山。而這樣的相似性,不僅限於某些特定的植物種類,而是遍及維管束植物的多個類群,顯然這種現象並不是特例,而是臺灣植物的一個重要的原鄉。
很多人在聽聞這本書之初,都會問一個問題:「這本書會很硬嗎」?誠然,這是本科普書,所以當然會有比較硬的知識性內容,不過在流暢的文字以及詳盡的照片輔助下,這些知識變得親民許多,就算只看書中精美的照片,仍能夠領會作者心心念念、如海一般廣大的山之國度。對於有在臺灣登山或是野外觀察的讀者,讀起來應該會感到莫名的熟悉,因為這本書寫的就是臺灣的山以及山上的植物,凡是登山都會遇見以及置身其中的那些風景。但是請小心,當你看完這本書之後,或許會不小心被橫斷山的山水以及高山花園推坑,陷入另一種登山中毒的症狀。
不過最後還是要說:作者應該可以處理一下我們的稿件了吧?

一個還是兩個科?杉科與柏科的分類之爭
杉科 (Taxodiaceae) 植物在目前的研究中,多被視為柏科 (Cupressaceae) 的一部分,對於很多習慣使用杉科的人來說,這或許是個難以接受的處理,因為過去教學中,兩個科有極大的差異,為何現在說併就併呢?兇手難道又是分子生物學研究的結果嗎?
在說明這個故事之前,我們得先回到1926年,這一年,恩格勒系統有關杉科與柏科的部分出版,這部分的著者是Pilger,而他撰寫的杉科與柏科的分類概念,就這樣被往後許多學者使用了數十年之久。從形態上看,兩者雌球花的苞鱗 (bract scale) 有相似的癒合狀態,所以主要的差別就在於營養器官上。杉科植物具有狹長且螺旋狀著生或互生的葉片;相對的,柏科植物具有鱗片狀且十字對生的葉片。不過這樣的區分方式還是有例外,例如杉科水杉屬 (Metasequoia) 植物具有對生的葉片,而產於塔斯馬尼亞的密葉杉屬 (Athrotaxis) 則具有鱗片狀的葉片。所以從形態上看來,兩者並不能非常有效的區分開來。


除了外觀相似之外,其他研究的結果也漸漸使人們懷疑杉科和柏科是否真正獨立。例如,Eckenwalder在1976年對各種針葉樹科進行形態和化學成分分析後,結果發現杉科和柏科之間的相似度高達90%,與其他科的相似度則只有29-59%。因此,他認為杉科和柏科應該合併為一個科,並建議一個新的分類系統。此外,Hart在1987年利用形態和化學成分資料進行支序學分析後, 發現杉科和柏科在演化樹上共同形成了一個有良好支持度的單系群。Price和Lowenstein在1989年也對種子蛋白質進行了免疫學研究,發現柏科和杉科在演化樹上形成一個與其他針葉樹類群明顯不同的分支,其中柏科和杉科還分別形成了5個不同的亞群,這表明杉科和柏科之間的差異是微小的。因此,這些研究結果表明,把杉科和柏科分為兩個獨立的科是不合適的。
這些研究都顯示一件事:杉科與柏科的界線並不如想像的涇渭分明,兩者之間有太多難以區分的部分,所以這樣的分類問題勢必需要修正。然而,上述的這些研究並非針對兩個科的劃界問題探討,在下進一步的決定之前,還需要更多的證據。
首先是來自於Brunsfeld的團隊在1994年的研究,這個團隊利用葉綠體的rbcL序列重建杉科與柏科的親緣關係,並且以南洋杉科 (Araucariaceae)、三尖杉科 (Cephalotaxaceae)、松科 (Pinaceae) 與羅漢松科 (Podocarpaceae) 等科植物為外群。結果顯示杉科和柏科共同形成一個支持度良好的單系群,其中柏科各屬亦共同形成良好的單系群,但杉科各屬則為並系群,所以這個結果亦支持兩者應該合併為一個科。Gadek等人也在2000年利用葉綠體的matK和rbcL序列,再度探討了這個議題,結果與Brunsfeld的研究結果相同,杉科仍然是個並系群,本研究不僅再次確認了這個事實,同時也根據研究結果,提出了廣義柏科 (Cupressaceae s.l.) 的新分類系統。
此後廣義柏科的概念逐漸被研究者接受,雖然從上述研究之後,仍然陸續有學者探討柏科的親緣關係,但是重點已經轉為探討柏科的親緣關係以及科內分類系統的調整,以及本科植物的起源與散布等議題 (例如Liu等人2022年的研究)。目前許多近期出版的圖鑑和資料也採用了這樣的分類概念,顯示廣義的柏科也正在進入到普及性的出版物中,成為植物學研究中至關重要的一個概念。此外,隨著分子生物學技術的發展,科學家們還可以進一步分析柏科與其他植物科別的遺傳序列,以更精確地瞭解它們之間的親緣關係,例如全基因組定序下的研究,或許又可以讓我們更加了解這群植物的關係,結果又可能再次改變我們對柏科植物的理解,而這就是分類學研究逐漸演進的痕跡與結果。
參考文獻:
Brunsfeld et al. 1994. Phylogenetic relationships among the genera of Taxodiaceae and Cupressaceae: evidence from rbcL sequences. https://doi.org/10.2307/2419600
Liu et al. 2022. Phylogeny and evolution of Cupressaceae: Updates on intergeneric relationships and new insights on ancient intergeneric hybridization. https://doi.org/10.1016/j.ympev.2022.107606

餐桌上的植物005—從米的結構出發,精通洗米
生活在台灣的人,米是主要的食物之一。除了直接煮成白飯,還有許多其他的料理方式。米在台灣人的生活中扮演著重要的角色,可以說是我們的血液中都充滿著米的DNA。因此,我們對於米的正統性也有著一種匠人般的堅持。幾年前有一部烹飪的YouTube影片示範烹煮米飯的方式是將米放入布袋中水煮。但這個方式遭到了網民的批評,認為這樣不符合傳統。可見,米對於我們來說是一種必須堅持正統方式的食材
很多人會把煮出好吃的白米飯視為評斷一個人廚藝的標準。如果在Google上搜尋如何煮好白米飯,會得到很多不同的方法。例如,添加油脂或香料,或是注重挑選米的品種等等,這些方法確實可以提高白飯的美味程度。但如果回到最基本的層面,我們常常在一開始就犯了錯誤。
首先讓我們回到米的原點,也就是米是怎麼從稻穀成為白米的。
我們吃的白米,其實是把水稻 (Oryza sativa) 的穎果 (caryopsis),其實主要是胚乳 (embryo) 的部分,其他不是胚乳的部分則隨著脫殼的過程一併被去除了。所以白米就像是削皮後的蘋果,是相當缺乏保護且脆弱的。因此,我們在賣場買到的白米,多半都是在真空的狀態,減少在搬運過程中的摩擦,以及米粒中水分的散失。
洗米是最重要的關鍵
很多煮飯的流程中,都會從洗米開始,各位應該有經驗,洗米的水通常會是深淺不定的乳白色,這些是什麼東西呢?上一段中提到米其實是胚乳,而它的主要成分是澱粉 (starch),也就是和玉米粉或太白粉類似的東西。所以在洗米的時候洗掉的,其實是米粒在相互摩擦或是保存過程中隨著水分散失崩解的澱粉。這些澱粉和白飯的外觀與口感息息相關,可以想像一件事,這些澱粉一但和米一起烹煮,這些澱粉在膨脹糊化之後,就會填補米粒間的空隙,造成白飯具有黏糊、令人不悅的口感。所以為了避免這種狀況,洗米的時候應該把這些澱粉盡量去除,也就是把米洗到流水清澈、米粒半透明的狀態,減少不必要澱粉的干擾。
另一個重點:米的保存
除了洗米之外,在米的保存過程中,減少米粒摩擦和崩解產生澱粉,也是讓米更好吃的關鍵。我們可以觀察一下賣場裡販賣的剛剛碾好的米,這些米通常看起來是呈現右上缺一角的橢圓形、而且是半透明的狀態,這就是米還充滿水分下的形態。然而在開封烹煮後,米的水分會不斷的散失,所以米粒的顏色會從半透明逐漸變白,形狀也會崩解成更小且不規則的狀態。這樣的變化會對白飯造成什麼影響呢?這些形狀和含水量不一的米粒在烹煮過程中的澱粉糊化程度不同,所以每一粒米的成熟度就不同,特別碎或小的米粒,會因為吸水過多造成過度軟爛,造成和上一段裡提到的澱粉粒同樣的效果。所以白米買回來之後盡可能保持密封和低溫,都有助於延長米的最佳狀態。
一碗好的白飯的口感與外觀,其實在保存和洗米的過程中就已經決定大半了。就算使用品質再好的米,如果保存方式不佳或清洗方式粗糙,也無法展現米本來的風味和口感。相反地,品質平庸的米可能因為良好的保存和仔細的清洗方式而表現得相當優秀。下次買米時,可以參考以上的小訣竅,讓你也能煮出令人滿意的一碗白飯。
傳說中水煮白飯的影片:
https://www.youtube.com/watch?v=KpKPueBqFWc

以福爾摩沙之名
說到第一個被發表的台灣植物種類的話,有許多研究都指出是由Hooker於1852年發表的蓪草 (Aralia papyrifera=Tetrapanax papyrifera)。那麼,如果換個問題,「第一個以台灣之名發表的植物」呢?
這個問題的答案,可以從線上資料庫的結果來告訴我們。根據搜尋的結果,第一個以Formosa為尾名的植物發表於1866年,係由時任英國外交官的Hance發表的翻白草 (Potentilla formosana=P. discolor)、楓香 (Liquidambar formosana)、紅子莢蒾 (Viburnum erosum var. formosanum=V. formosanum) 以及台灣石楠 (Photinia villosa var. formosana=P. lucida)。這四種植物都由Oldham採自淡水附近,為海岸或淺山地區的植物。其中翻白草早已消失於淡水地區,也算是一個歷史的見證吧!
如果是Taiwan的話,那結果又是什麼呢?相當出人意料的,最早以Taiwan當做尾名的是台灣溲疏 (Deutzia crenata var. taiwanensis=D. taiwanensis),由俄國植物學家Maximowicz發表於1867年的一篇東亞的八仙花科植物專論中。有趣的是,作者引證的標本同樣是Oldham採自淡水附近,這是巧合嗎?我不知道,但有可能。順帶一提,在種階最早用Taiwan作為尾名的是赫赫有名的台灣蘇鐵 (Cycas taiwaniana),由Carruthers發表於1893年。
最後有一個有趣的問題要考考大家,地名後綴-ensis或-ense與屬名的性別有關,前者用於陽性和陰性屬名,而後者用於中性屬名。問題來囉,台灣唯一一個以formosense發表於種位階的學名,是哪一個呢 (因轉移產生的種尾名變化不計的話)?而發表於變種階的又是哪一個呢?
本文最初於2021.1.13刊登於同名專頁

2020—台灣維管束植物的新成員

為什麼SOP在指定模式標本上不可行?
2017年在深圳舉辦的國際植物學大會中,有關法規的討論中最主要且影響最大的變革,就是和所謂的美國法規 (American code) 或費城法規 (Philadelphia code) 裡提出的一種指定屬或屬下類群 (不包括種) 的方法有關。
如果對植物命名法規的歷史稍有了解的話,應該會有印象因為對維也納會議 (1905年) 產生的結果不滿的緣故,美國學者提出了另一個版本的法規,也就是美國法規 (1907年),其前身即為1904年提出的費城法規。在這兩個法規當中,都對指定屬或屬下類群的方式,提出一種標準化的做法,也就是在深圳法規所謂的「largely mechenical method (大量機械法)」。這個方法被記錄於美國和費城法規中的Canon 15: 屬或屬下類群的模式是作者命名所根據的那個種,或作者另行指定的種。若無上述指定,則為該類群中根據下列規則的第一個種 (下略)。
從現在的角度看來,這樣的規定與法規的宗旨背道而馳,但當時仍有一些學者根據該規定指定的很多的模式,因此IAPT成立了一個特殊的委員會 (Special Committee on Publications Using a Largely Mechanical Method of Selection of Types) 來處理有關這個規定指定的模式問題。
在深圳法規的規定中,有關大量機械化的規定主要在10.5, 10.6和10.7條款。10.5條款規範直接說明使用該方法選擇的模式可以被不使用該方法指定的模式取代。10.6條款則定義何謂大量機械化法 (當然主要是美國和費城法規)。10.7條款則說明1935年1月1日前的文獻中,如何認定該文獻作者是否採用該方法指定模式。
至於為何是1935年1月1日呢?如果翻開法規的歷史會發現,1935年的劍橋法規 (根據1930年的劍橋會議) 調和了維也納法規和美國法規之間的差異,也解決了植物學上派系爭議,使法規真正在國際上普及化。
有關費城法規和美國法規的法條內容可以參考以下文獻:Bulletin of the Torrey Botanical Club 31(5):249-261, 1904 (費城法規). 相同期刊的34(4):167-178, 1907 (美國法規)。
有關特別委員會的相關報告可以參考Taxon 65(6):1441-1442和1443-1448, 2016。
本文最初於2020.8.27刊登於同名專頁

拍攝花粉的know how
前面討論了幾次有關花粉的形態以及前處理方式的差異及注意事項,花粉系列的最後一彈就是要解析如何有效的取得花粉的各種特徵。由於近年來掃描式電子顯微鏡 (SEM) 的操作門檻降低的緣故,使用該技術觀察花粉變成主流的觀察方式,因此以下的說明也是根據該技術而來。
如同其他研究一樣,首先要做的事就是收集文獻,然後分析這些文獻使用的方法和拍攝成果,例如該研究使用酸解法,那麼該研究的花粉是否有外壁塌陷的現象 (通常不會很明顯,但有的時候可以看到如餡餅狀壓扁的花粉),或是使用酒精序列脫水,那麼可以觀察是否有明顯的花粉鞘。如果該研究是直接用烘乾標本上的花粉,那麼可以預期該花粉是處於脫水且溝孔關閉的狀態。這些研究有助於判斷前處理的方式,例如花粉鞘較少甚至缺少的狀態,可以使用酒精保存花粉,若花粉在調和作用前後變異不大,或是沒有變異,那麼甚至可以直接烘乾,不需要經過序列脫水和臨界點乾燥。
其次是花粉的前處理,考慮到花粉的調和作用和花粉鞘,花粉可以分成直接乾燥和浸液兩組,其中後者建議直接使用調配好的DMP溶液,可以有效的脫水和去除花粉鞘。需要注意的是,任何前處理液都無法保證花粉的膨潤狀態,因此在浸泡之前須確定花粉是處於膨潤的狀態,必要的話可以泡水再浸泡至DMP。
另外在拍攝的時候,建議由大至小,先拍極面和赤道面的全景,然後再拍兩個面的雕紋。如果花粉有溝孔,那麼溝孔中的雕紋也要紀錄。另外如果花粉表面有其他的附屬物,也要一併紀錄。以上的紀錄步驟在兩組分別處理的花粉中要分別紀錄,如果要加上後續量測用的數量,那單一樣本要拍的照片數量就會很多,建議使用系統化的表格,以免之後還要補。此外,花粉的照片在後續常需要製作成圖版,因此最好稍微紀錄一下拍攝的角度和比例尺,方便之排版作業。
此外,有些研究會關注花粉內部的形態,通常是探討花粉壁的結構。觀察這類特徵的方式通常是利用穿透式電子顯微鏡 (TEM),將花粉經過包埋切片後拍攝,缺點是前處理的流程較長,取得的影像缺乏立體感等。另一種較少見的方式是利用冷凍切片加上SEM拍攝取得花粉剖面的影像,這個方法雖然可以取得相對有立體感的影像,但前處理跟一般SEM的流程相比複雜許多,有興趣的話可以參考Vezey et al. (1994) 的研究。
本文最初於2020.8.14刊登於同名專頁